Le système digestif des Ruminants, et notamment leur « estomac », présente un certain nombre de particularités leur permettant de digérer les végétaux qu'ils ingèrent. Après avoir présenté ces spécificités, cet article explique le rôle du microbiote ruminal dans la digestion des glucides, lipides et protéines.
Les Ruminants possèdent trois réservoirs prégastriques dont le plus important est le rumen. Il fait jusqu’à 200 L chez un bovin adulte et autour de 15 L chez un ovin, et permet la digestion microbienne des aliments, en particulier ceux d’origine végétale. Le bon fonctionnement du rumen nécessite des conditions particulières : une température de 39 °C, un pH voisin de 6 et une anaérobiose stricte.
Les digestions ruminales sont réalisées par des Bactéries, des protozoaires, des Archées et des Champignons. Ces microorganismes dégradent les glucides en acides gras volatils, transforment une partie des protéines ingérées, et hydrolysent les triglycérides et autres esters, puis assurent l’hydrogénation de la majorité des acides gras insaturés (Figure 1).
Selon la nature du glucide fermenté, on différencie deux types de bactéries. Les bactéries amylolytiques, dont le substrat privilégié est l’amidon, synthétisent essentiellement du propionate et préfèrent les pH inférieurs à 6. Les bactéries fibrolytiques s'attaquent quant à elles surtout aux glucides pariétaux pour donner majoritairement de l’acétate et du butyrate, et affectionnent les pH supérieurs à 6.
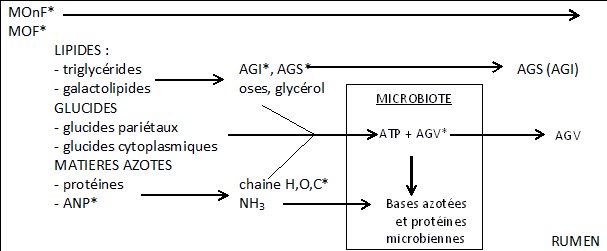
MOnF : matière organique non fermentée, MOF : matière organique fermentée, ANP : azote non protéique, AGS : acides gras saturés, AGI : acides gras insaturés, AGV : acides gras volatils.
Le rumen et son écosystème microbien
Particularité anatomophysiologique du tube digestif des Ruminants
Anatomie de l’estomac des Ruminants
Les Ruminants possèdent trois pré-estomacs : le réseau et le rumen, formant le réticulorumen, et le feuillet. Le « véritable » estomac, sécrétant des sucs gastriques, est la caillette. Cet ensemble est très volumineux : il représente environ les deux tiers de la cavité abdominale.
| Noms | Aspect de la paroi | |
|---|---|---|
| Panse | Rumen | Papilles |
| Bonnet | Réticulum ou réseau | Alvéoles polygonales |
| Feuillet | Omasum | Lames parallèles |
| Caillette | Abomasum | Muqueuse sécrétrice |
Le réseau est le plus petit et le plus antérieur des pré-estomacs. L’œsophage est abouché dorsalement au réseau. Ce dernier est prolongé dorsalement par le rumen (ou panse). Le rumen est un vaste sac bilobé, allongé d’avant en arrière. La muqueuse du rumen est couverte de papilles (de 2 mm à 2 cm), ces papilles sont kératinisées mais l’épithélium est fin et très vascularisé. Les papilles ont pour principal rôle d’absorber les produits des fermentations microbiennes.
L’omasum (ou feuillet) est le dernier pré-estomac. Il est placé entre le rumen et la caillette. Le feuillet est presque entièrement occupé par des lames parallèles, de hauteurs inégales, disposées dans le sens du transit alimentaire, et dont la muqueuse est de même nature que celle du rumen. Elle correspond à une surface d’absorption additionnelle.
L’abomasum (ou caillette) est comparable à l’estomac des monogastriques.
Le reste du tube digestif est comparable à celui des monogastriques. On retrouve l’intestin grêle et le gros intestin. Cet ensemble mesure environ 50 m chez le bovin adulte. Les trois portions de l’intestin grêle sont le duodénum, le jéjunum et l’iléon. Le gros intestin est composé du cæcum, du côlon et du rectum.
La motricité des estomacs des Ruminants
Le rumen est un fermenteur anaérobie où la digestion microbienne se déroule en continu. Les mouvements de ce réservoir brassent la masse alimentaire et facilitent son ensemencement bactérien. Ces mouvements participent également à la régurgitation physiologique du bol alimentaire qui va pouvoir être à nouveau mastiqué et insalivé : c’est la rumination. Ensuite, ils permettent la vidange vers l’omasum. Puis ils contribuent à l’élimination des gaz de fermentation : c’est l’éructation.
Ruminants et ruminants
Les ruminants sont des herbivores qui ingèrent en peu de temps une grande quantité de nourriture en la mâchant peu. Le bol alimentaire est rapidement envoyé dans l’« estomac » de l’animal. Plus tard, le contenu de l’estomac est régurgité pour être longuement mâché, ce qui permet de fragmenter la nourriture et donc d’augmenter la surface de contact entre celle-ci et les microorganismes et enzymes responsables de la digestion.
La rumination est apparue indépendamment à plusieurs reprises au cours de l’évolution. On la retrouve ainsi chez les Tylopodes (dromadaires, chameaux, guanacos, vigognes…), les Kangourous et les Ruminants (bovins, ovins et caprins notamment).
Les conditions physicochimiques dans le rumen
Le rumen a un volume moyen d’environ 150 L, dont 90 L de digesta chez les bovins (4). Ce contenu n’est pas réparti de façon homogène dans le rumen : en partie ventrale on trouve une phase liquide, en partie intermédiaire une phase solide et en partie dorsale une phase gazeuse.
La phase liquide a pour origine l’abreuvement (50 à 100 L par jour), la salivation (80 à 200 L par jour) et l’eau contenue dans les aliments. L’eau est le constituant principal du contenu ruminal (85 %), et se trouve principalement dans la phase liquide contenant de fines particules en suspension (particules alimentaires ou bactéries) et des molécules en solution (sels minéraux, petites molécules organiques). Cette phase liquide permet l’imbibition des aliments. L’eau est essentielle aux réactions réalisées par les enzymes microbiennes.
La phase solide se concentre dans un amas fibreux en partie dorsale du rumen et a pour origine l’ingestion d’aliments. Quant à la phase gazeuse elle comprend majoritairement les gaz issus des fermentations microbiennes. Ces gaz sont éliminés par éructation.
Le pH ruminal est normalement compris entre 5,5 et 7. Ce pH est la résultante d’un équilibre entre des acides faibles et des bases faibles, issus des fermentations microbiennes et des substances tampons apportées par la salive (bicarbonates et phosphates) (5).
La température du rumen est comprise entre 39 et 41 °C, soit environ 1 °C au-dessus de la température corporelle. Elle augmente en fonction de l’intensité des fermentations ruminales et est donc maximale dans les heures suivant le repas.
Le rumen abrite donc des conditions physicochimiques très particulières permettant le développement d’un microbiote anaérobie très actif.
Le microbiote
Le rumen accueille un microbiote diversifié : environ 200 espèces de bactéries (1010 à 1011 bactéries par mL), des protozoaires (de 104 à 106 par mL) et des champignons (entre 103 et 105 zoospores par mL – les zoospores sont des spores mobiles flagellées participant à la reproduction de certains Eumycètes). On trouve également entre 107 et 109 particules de virus bactériophages par mL (5).
Les Bactéries
Les Bactéries correspondent à un peu plus de la moitié de la biomasse microbienne du rumen. Les trois quarts de ces Bactéries sont fixés sur des particules alimentaires. Elles sont constamment éliminées par prédation des protozoaires ruminaux ou évacués vers le feuillet et le reste du tube digestif. Néanmoins, ces pertes sont compensées par une croissance régulière de la population bactérienne.
Cette population est de loin la plus active. Les Bactéries du rumen sont généralement classées selon leur capacité à dégrader certains substrats et à les utiliser pour leur survie. En particulier elles sont souvent distinguées en fonction de leur activité glucidolytique : fibrolytique ou amylolytique.
Les principales espèces fibrolytiques sont Fibrobacter succinogenes, Ruminococcus albus, Ruminococcus flavefaciens et Butyrivibrio fibrisolvens. Ces Bactéries adhèrent aux particules fibreuses et synthétisent des enzymes actives sur les glucides pariétaux (cellulose, hémicelluloses et pectines). Cette population se développe mieux avec un pH supérieur ou égal à 6,5. Les principales espèces amylolytiques (digérant l’amidon) sont Streptococcus bovis, Ruminobacter amylophilus, Succinomonas amylolytica, Selenomonas ruminantium et Prevotella ruminicola. Elles préfèrent des pH inférieurs à 6.
Il n’existe pas de bactérie strictement protéolytique : ce sont en grande partie les Bactéries amylolytiques (et certaines Bactéries fibrolytiques) qui exercent cette fonction.
Les lipides sont soumis à deux types de réactions qui s’enchaînent : la lipolyse des esters d’acides gras suivie par la biohydrogénation des acides gras insaturés.
La lipolyse est principalement réalisée par Anaerovibrio lipolytica, et certains représentants du genre Butyrivibrio.
La biohydrogénation ruminale est complexe : elle se divise en plusieurs étapes successives d’isomérisations et de réductions et elle nécessite l’intervention de plusieurs enzymes synthétisées par des espèces différentes. Les Bactéries impliquées dans ce phénomène ne sont pas encore très bien connues si ce n’est celles appartenant au genre Butyrivibrio.
Les protozoaires
Les protozoaires ruminaux (comme Endotinium caudatum ou Epidinum ecaudatum) sont des organismes eucaryotes unicellulaires microscopiques. Ils sont de taille variable, 20 à 100 fois plus grands que les Bactéries mais 104 fois moins nombreux. Ils peuvent représenter jusqu’à 40 % de la biomasse microbienne. La plupart des protozoaires sécrètent des enzymes (protéolytiques, fibrolytiques, amylolytiques) participant à la digestion des particules ingérées (4).
Les protozoaires sont des prédateurs des Bactéries et récupèrent les acides aminés, les peptides et les acides nucléiques bactériens pour les incorporer dans leur propre organisme (4).
Les Champignons
Les Champignons appartenant principalement aux genres Neocallimastix, Piromyces et Caecomyces, sécrètent de nombreuses enzymes intervenant dans la digestion des glucides pariétaux, mais ont un rôle mineur par rapport aux bactéries. Ils ont une activité protéolytique faible. Leur contribution à la dégradation des glucides cytoplasmiques est peu connue.
Les Archées
Au sein du microbiote ruminal, figurent également des Archées méthanogènes qui utilisent le dihydrogène produit par le métabolisme bactérien en conditions anaérobies pour réduire le dioxyde de carbone en méthane. Cette réaction est nécessaire car la présence en excès de dihydrogène inhiberait les fermentations et donc le fonctionnement du rumen. Dans le gros intestin ce sont principalement les Bactéries acétogènes qui remplissent cette fonction. Les Archées ne participent pas directement à la digestion ruminale. Elles forment une population peu abondante et probablement peu diversifiée et encore peu connue, mais qui serait peu diversifiée.
Le rumen est donc un fermenteur, présentant des conditions physicochimiques particulières, qui permet la multiplication d’un microbiote anaérobie. Il permet ainsi aux Ruminants de tirer parti des fibres végétales. Nous allons maintenant nous intéresser aux processus de dégradation des principaux composants alimentaires.
Digestion des aliments d’origine végétale par les microorganismes ruminaux
Digestion des glucides
Les glucides des végétaux
Les glucides peuvent se classer en deux catégories selon leur répartition dans la cellule végétale. La première catégorie correspond aux glucides cytoplasmiques : ils sont contenus à l’intérieur de la cellule végétale. Ce sont des substrats ou des intermédiaires du métabolisme cellulaire des glucides, et des glucides de réserve. Les glucides cytoplasmiques comprennent donc les glucides hydrosolubles (fructose, saccharose, glucose…) et les polyholosides de réserve (amidon).
La seconde correspond aux glucides pariétaux : on ne les retrouve que dans les aliments d’origine végétale ; ils sont, avec la lignine, polymère phénolique, les constituants de la paroi végétale : cellulose, hémicellulose, pectines. Les hémicelluloses et la cellulose sont systématiquement liées et enchevêtrées pour constituer la base de la paroi végétale. Les hémicelluloses sont moins résistantes à l’hydrolyse que la cellulose et elles ont une composition beaucoup plus variée. Les pectines forment l’ensemble des fibres dites solubles.
Les glucides pariétaux ne peuvent pas être hydrolysés par les enzymes glycolytiques sécrétées par le tube digestif. En effet ces dernières ont une action α-glycosidase mais non β-glycosidase et ne sont pas capables d’hydrolyser les liaisons assemblant les polymères d’acide α-galacturonique composant les pectines. Cependant, les glucides pariétaux peuvent être dégradés par les enzymes sécrétées par le microbiote ruminal, capables de catalyser ces réactions. Ces différences s’expliquent par la structure chimique et l’encombrement stérique des polyosides considérés. En effet, si la liaison se réalise entre les mêmes fonctions des mêmes composés, il existe une différence d’encombrement stérique entre la liaison α-osidique et la liaison β-osidique. Cette modification stérique est à l’origine d’une inadéquation entre le site catalytique très spécifique des enzymes à action α-glycosidase et la liaison β.
La dégradation extracellulaire des glucides
Les bactéries fibrolytiques sont les principales effectrices de la dégradation des glucides pariétaux, mais les champignons (comme Neocallimastix frontalis) et certains protozoaires (comme Entodinium caudatum) peuvent être impliqués. L’action de ces bactéries est facilitée par un pH supérieur à 6,5. Les bactéries vont tout d’abord se fixer aux fibres alimentaires. Certains protozoaires font de même, cependant, en général, ils ingèrent des petites particules alimentaires en suspension. Les champignons utilisent leurs rhizoïdes (fins filaments de mycélium ressemblant à des racines, et utilisés pour la fixation du champignon à son substrat) pour pénétrer en profondeur les fragments végétaux (4) ; ils coloniseraient préférentiellement les tissus lignifiés (6).
Les bactéries sécrètent de nombreuses cellulases dans le milieu ruminal, dégradant la cellulose en cellobiose et en cello-oligosaccharides de faible degré de polymérisation puis en glucose (4), et des hémicellulases pour dégrader les hémicelluloses (6) (Figure 3). Les protozoaires et les champignons sécrètent également des hémicellulases et des cellulases (6). Pour ce qui est de l’hydrolyse des pectines, des enzymes pectinolytiques ont été identifiées chez des bactéries et des protozoaires mais pas chez les champignons (6).
Les bactéries amylolytiques et les protozoaires sont impliqués dans l’hydrolyse de l’amidon. Les bactéries amylolytiques sont favorisées par un pH ruminal inférieur à 6. Les microorganismes sécrètent des amylases extracellulaires, qui vont hydrolyser l’amidon. Le produit final de cette hydrolyse est le glucose.
L’hydrolyse extracellulaire des glucides végétaux complexes produit des glucides simples qui vont pouvoir être absorbés et fermentés par les microorganismes, principalement les bactéries.
La fermentation des glucides par les bactéries ruminales
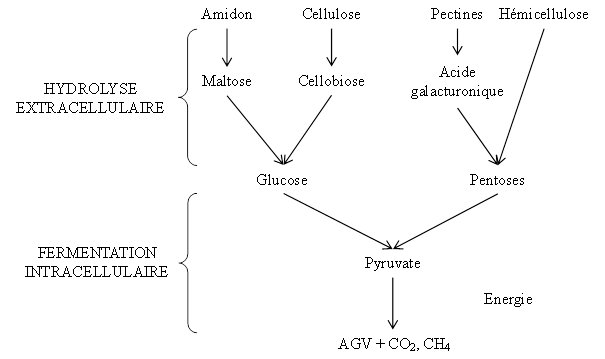
Le glucose et les pentoses issus de la digestion extracellulaire sont absorbés par les bactéries. Le pyruvate est le premier intermédiaire formé et est rapidement utilisé. Il est catabolisé en acides gras volatils (AGV), en libérant des gaz (dioxyde de carbone et méthane). Cette réaction est couplée à la formation d’ATP qui permettra le métabolisme et la croissance microbiens (6). Les deux étapes de la dégradation des glucides sont illustrées dans la figure 3. Les AGV sont absorbés au niveau de la muqueuse ruminale.
Digestion des lipides alimentaires
Les différents lipides alimentaires
Les rations des Ruminants sont généralement pauvres en lipides. Les lipides des grains de céréales, des graines protéagineuses et oléagineuses sont principalement composés de triglycérides riches en acides gras insaturés : acide α-linolénique (C18:3-ω3), acide linoléique (C18:2-ω6) et acide oléique (C18:1-ω9).
L’herbe contient essentiellement des galactolipides et des phospholipides qu’on retrouve principalement dans les feuilles. Les acides gras majoritaires de l’herbe sont le C18:3-ω3, le C18:2-ω6 et l’acide palmitique (C16:0) qui représentent respectivement 41, 19 et 24 % des acides gras totaux de l’herbe (7,8).
Les acides gras
Les acides gras sont des acides carboxyliques possédant une chaîne carbonée plus ou moins longue. Cette chaîne carbonée peut contenir des doubles liaisons carbone-carbone, auquel cas l’acide gras est qualifié d’insaturé. Si par contre la chaîne carbonée ne contient que des liaisons simples carbone-carbone, l’acide gras est dit saturé (car on ne peut pas « ajouter » plus d’hydrogène à la molécule).
La notation Cx:y permet de rendre compte de la nature de l’acide gras : x représente le nombre d’atomes de carbone et y le nombre de doubles liaisons.
Le métabolisme ruminal des lipides
Le métabolisme ruminal des lipides est effectué en deux étapes : la lipolyse, suivie de la biohydrogénation. Cette deuxième étape entraîne le remaniement des AG alimentaires et donc la production de nombreux isomères de ces AG. De plus les bactéries ont la capacité de synthétiser de nouveaux AG. L’ensemble de ces phénomènes est à l’origine d’une composition très variée d’AG ruminaux (6).
Les triglycérides, les galactolipides et les phospholipides sont tout d’abord rapidement hydrolysés par les bactéries lipolytiques en milieu extracellulaire : cela donne des acides gras libres, du glycérol, des oses et du phosphate. Cette hydrolyse est quasi complète (entre 85 et 95 %) et rapide. Les acides gras insaturés vont ensuite subir la biohydrogénation tandis que le glycérol et les oses seront fermentés en AGV.
La biohydrogénation est une réaction extracellulaire réalisée par diverses enzymes produites par de multiples espèces bactériennes. Elle se déroule en plusieurs étapes qui vont saturer progressivement les acides gras insaturés.
La biohydrogénation commence par une réaction d’isomérisation. Les premiers intermédiaires formés présentent deux doubles liaisons conjuguées (c’est-à-dire séparées par une seule liaison simple). Ces intermédiaires sont appelés acides linoléniques conjugués (CLnA) et acides linoléiques conjugués (CLA) respectivement pour le C18:3-ω3 et le C18:2-ω6. Ces intermédiaires auraient des propriétés biologiques intéressantes pour le consommateur humain, en particulier les CLA (9). La deuxième phase de la biohydrogénation correspond à l’hydrogénation des doubles liaisons par des réductases microbiennes produisant de nombreux intermédiaires et dont le produit final est l’acide stéarique (C18:0).
Cette biohydrogénation étant plus ou moins complète, il est possible de retrouver tous les intermédiaires de la biohydrogénation dans le contenu ruminal. Tous ces acides gras seront absorbés dans l’intestin grêle, et peuvent se retrouver dans le lait ou la viande des animaux. Elle est à l’origine d’une teneur plus grande en acides gras saturés des produits de Ruminants.
Digestion des protéines alimentaires
Nature de l’azote alimentaire
Les aliments d’origine végétale les plus riches en protéines sont les graines oléagineuses et protéagineuses, et certains coproduits de l’industrie agroalimentaire de ces graines et des céréales. Dans les plantes fourragères, 75 à 85 % de l’azote est sous forme de protéines, 15 à 25 % sous forme d’azote non protéique (ANP). L’ANP regroupe des amides et des acides aminés libres (50 % environ), ainsi que des bases azotées, des amines, de l’ammonium (NH4+), de l’urée et des nitrates (NO3-). Les protéines se concentrent dans les feuilles alors que l’ANP est surtout présent dans les tiges (10).
Les Mammifères éliminent l’azote excédentaire sous forme d’urée dans l’urine. Chez les Ruminants cette molécule peut aussi être excrétée dans la salive. Ce recyclage permet d’apporter aux microorganismes de l’estomac des Ruminants l’azote nécessaire à leur fonctionnement et de compenser ainsi en partie une faible teneur de ce composé dans les aliments ingérés. Dans les élevages, de l’urée peut être ajoutée à la ration alimentaire des animaux.
Dégradation des protéines
La protéolyse est principalement réalisée par les bactéries et les protozoaires. Les champignons interviennent dans une moindre mesure. Ces microorganismes ruminaux dégradent une fraction des protéines alimentaires. Cette dégradation engendre tout d’abord des peptides puis des acides aminés et enfin de l’ammonium. L’ANP est quant à lui rapidement dégradé en ammonium.
La première étape de la dégradation des protéines résulte de l’activité extracellulaire des protéases bactériennes. Ce sont essentiellement les bactéries amylolytiques qui interviennent dans la protéolyse : elles dégradent principalement les protéines solubles. Les protozoaires participent plutôt à la dégradation des protéines insolubles incluses dans des particules de grande taille telles que les chloroplastes ou les bactéries qu’ils ingèrent (11).
La vitesse de dégradation des protéines dépend tout d’abord de la solubilité : les protéines insolubles sont plus lentement dégradées que les protéines solubles. Il est également essentiel que les protéines soient accessibles : de nombreuses protéines végétales sont incluses dans une matrice de fibres. La protéolyse dépend alors de l’activité fibrolytique. Ainsi une fraction variable de protéines échappe à la dégradation ruminale. Elle se compose principalement de protéines insolubles et est représentée par les protéines alimentaires sur la figure 1.
Cette première hydrolyse extracellulaire par des enzymes bactériennes produit majoritairement des acides aminés, qui sont ensuite absorbés par les bactéries. Les acides aminés, d’origine alimentaire ou produits de l’hydrolyse des protéines, sont soit incorporés directement dans les protéines bactériennes, soit désaminés. Le squelette hydrogénocarboné de l’acide aminé servira alors à la production d’AGV ou de nouveaux acides aminés dans la bactérie.
La synthèse microbienne des protéines
Seule une petite fraction des acides aminés est directement incorporée dans les protéines microbiennes (chez les protozoaires et quelques espèces bactériennes). La plus grande partie des acides aminés bactériens est synthétisée à partir de l’ammonium. Les bactéries l’incorporent à une chaine hydrogénocarbonée pour former un acide aminé. Ces derniers permettront la synthèse de protéines.
La croissance des bactéries non protéolytiques dépend donc de la production d’ammonium par les bactéries protéolytiques. Il s’agit là d’une illustration des interdépendances nutritionnelles qui permettent le bon fonctionnement du microbiote ruminal. Notons que les bactéries fibrolytiques sont très dépendantes car elles ont majoritairement une action protéolytique faible.
Les protéines d’origine microbienne ainsi que les protéines alimentaires non dégradées dans le rumen seront ensuite évacuées vers le reste du tube digestif. Leur digestion, dans le reste du tube digestif, est similaire à celle rencontrée chez les monogastriques.
Conclusion
Les activités ruminales permettent aux Ruminants de valoriser efficacement l’énergie des glucides pariétaux contenus dans les végétaux. Elles contribuent également à un remaniement des matières azotées présentes dans la ration, et rendent les Ruminants capables d’utiliser l’azote non protéique. Les protéines issues des synthèses microbiennes présentent des compositions en acides aminés globalement équilibrées : les activités microbiennes augmentent ainsi la valeur biologique des matières azotées alimentaires. Ainsi les Ruminants sont peu dépendants des équilibres entre les différents acides aminés alimentaires.
De plus, il est important de noter que les synthèses microbiennes à partir de l’ammonium nécessitent de l’énergie sous forme d’ATP, énergie provenant directement de la fermentation des glucides. Ces deux métabolismes sont donc étroitement liés.
Le métabolisme ruminal des lipides quant à lui, affecte grandement le profil en acides gras ingérés par l’animal. Il est à l’origine de la mauvaise image des produits de Ruminants, notamment les produits laitiers, car il enrichit les productions en acides gras saturés. Néanmoins il est aussi à l’origine de la formation d’acides gras particuliers et qui auraient de potentiels effets bénéfiques pour la santé des consommateurs, tels les CLA.
Références
- Sautet J. (1995). L’appareil digestif et ses adaptations. In : Nutrition des ruminants domestiques. INRA Editions, p 183-222.
- Thivend P., Fonty G., Jouany J.P., Durand M., Gouet Ph. (1985). Le fermenteur rumen. Reprod. Nutr. Dévelop., 25, 729-753
- Malbert C.H., Ruckebusch Y., Bueno L., Baumont R., Theodorou V., Brikas P. (1995). Motricité du complexe gastrique. In : Nutrition des ruminants domestiques. INRA Editions, p 223-252.
- Jouany J.P. (1994). Les fermentations dans le rumen et leur optimisation. INRA Prod. Anim., 7, 207-225.
- Dusart C. (2014). La digestion ruminale : mise en place d’un modèle d’étude in vitro à long terme en cultures de type « batch ». Thèse de doctorat vétérinaire
- Grenet E., Besle J .M. (1991). Microbes and fibre degradation. In : Rumen microbial metabolism and ruminant digestion. INRA Editions, p 107-129.
- Cuvelier C., Cabaraux J.F., Dufrasne I., Istasse L., Hornick J.L. (2005). Production, digestion et absorption des acides gras chez le ruminant. Ann. Méd. Vét., 149, 49-59.
- Doreau M., Fievez V., Troegler-Meynadier A., Glasser F. (2012). Métabolisme ruminal et digestion des acides gras longs chez le ruminant : le point des connaissances récentes. INRA Prod. Anim., 25 (4), 361-374.
- Troegeler-Meynadier A., Enjalbert F. (2005). Les acides linoléiques conjugués : 1. Intérêts biologiques en nutrition. Revue Méd. Vét., 156, 207-216.
- Jarrige R., Grenet E., Demarquilly C., Besle J.M. (1995). Les constituants de l’appareil végétatif des plantes fourragères. In : Nutrition des ruminants domestiques. INRA Editions, p 25-81.
- Jouany J.P., Broudiscou L., Prins R.A., Komisarczuk-Bony S. (1995). Métabolisme et nutrition de la population microbienne du rumen. In : Nutrition des ruminants domestiques. INRA Editions, p 350-381.



